Three Cognitive Computational Neuroscience Essays
The coursework for my Cognitive Computational Neuroscience (CCN) class this semester, was to write three essays. I was ~decently happy with my essays, so I will publish them here too.
Each of these essays were prompted - by the lecturer - with an essay topic statement, asking us to take a position on the given topic. Therefore, for each of these, I will first provide the topic statement (and position), and afterwards my essay will follow. It will be clear from the essays that, one some of the topics, I disagreed even with how the positions on the topic was presented, and discussed the question on more of a meta-level.
OBS: The pictures are now fixed to show the visual abstracts for each essay.
Essay 1
Essay topic: Modern convolutional neural networks [have/do not have] sufficient biological fidelity to be a good model for the brain
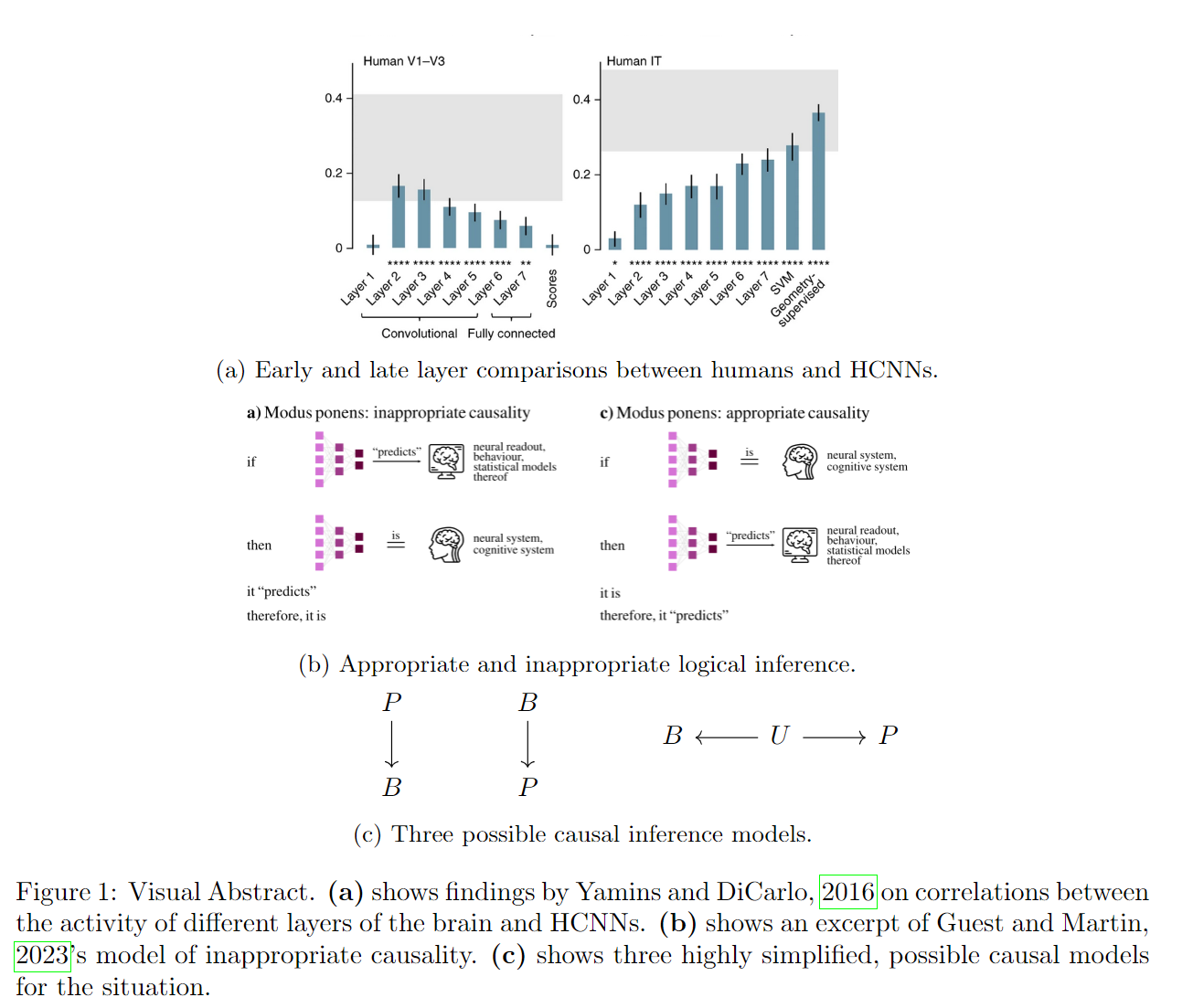
This essay engages with the topic on two levels: on one level by refuting the need for biological fidelity, and on another by discussing the need for causal inference. It starts by showing some empirical evidence for similarity between brains and Hierarchical Convolutional Neural Networks (HCNNs), then moves on to discuss whether we are committing a causal and logical fallacy when interpreting such data. After this follows a discussion on the use of causal inference diagrams. In their 2016 paper, Yamins and DiCarlo (2016) relay evidence showing that HCNNs that perform better at object recognition also do better at predicting cortical spiking data. The most striking finding comes from Khaligh-Razavi and Kriegeskorte (2014), who show that later layers of an HCNN are better at predicting the inferior temporal cortex than earlier layers, and likewise earlier layers the human V1-V3 areas, as shown in subfigure (a). This proceduro-structural simi-larity is solid evidence towards favouring the idea that HCNNs can be a good model for (parts of) the brain. Note that this does not require any level of biological fidelity. Finding the right sort of likeness, at the right level of analysis (Marr, 1982), will provide useful insight into solving the sorts of problems we care about. However, as Guest and Martin (2023) shows, we should be careful about what kinds of inferences we draw from our empirical observations. They place folk rules of inference about NNs into a formal system, and conclude that the inference ”if the neural network predicts brain data, then the neural network is a good model for the brain” is a case of inappropriate logical reasoning (subfigure (b)). They further argue, by bringing in the greater theoretical CCN scientific context, that the correct logical reasoning would be the converse: ”if it is a good model for the brain, then it predicts brain data.” The last missing piece is that both description and prediction should be placed within a context of causal inference. If the causal interplay between the HCNNs’ task performance and brain-likeness had been formalized in such a way, it would be open to critique, interpretation, and further theory. While papers often discuss these relationships informally, they do not appear to apply a rigorous causal inference analysis. In subfigure (c) B is the property of being Brain-like, P is Performance, and U a possible unobserved variable. The three causal inference diagrams describe causal relationships where, performance causes systems to be brain-like, being brain-like causes performance, and that some unobserved variable causes both performance and brain-likeness, respectively. Current statistical data can be equally well fit to either of these models; an actual intervention is needed to distinguish between these possible realities (Pearl et al., 2021, Chapter 3). This essay finds that efforts to analogize between neural networks and the brain are misguided in two ways. First, the focus on ”biological fidelity” is a possible distractor from discovering useful and generalizable properties of cognitive systems at different levels of analysis. Second, the lack of causal inference work in this field leads to a situation of confusion about where to proceed from, and what to do with, statistical relationships in data. Without an explicit causal model, to say that an HCNN is a model for the brain is to present a prediction without proper backing.
References
Guest, O., & Martin, A. E. (2023). On Logical Inference over Brains, Behaviour, and Artificial Neural Networks. Computational Brain & Behavior, 6 (2), 213–227. https://doi.org/10. 1007/s42113-022-00166-x Khaligh-Razavi, S.-M., & Kriegeskorte, N. (2014). Deep Supervised, but Not Unsupervised, Models May Explain IT Cortical Representation. PLOS Computational Biology, 10 (11), e1003915. https://doi.org/10.1371/journal.pcbi.1003915 Marr, D. (1982). Vision: A computational investigation into the human representation and processing of visual information. W.H. Freeman. Pearl, J., Glymour, M., & Jewell, N. P. (2021). Causal inference in statistics: A primer (Reprinted with revisions). Wiley. Yamins, D. L. K., & DiCarlo, J. J. (2016). Using goal-driven deep learning models to understand sensory cortex. Nature Neuroscience, 19 (3), 356–365. https://doi.org/10.1038/nn.4244
Essay 2
Essay topic: To fully reproduce the capabilities of the brain, we might need to also reproduce how activity changes over time.
While change in activity over time is the clearest indicator of what the brain is doing differently, we have to be careful about which features we think are necessary and/or sufficient for reproducing the brain’s capabilities. There are at least two features that seem important in this case: 1) biological neurons are connected in such a way to create loops and other feedback mechanisms (“recurrency”), 2) the brain adaptively changes in response to environmental and/or task demands. If we could recreate either of these \textit{without} implementing “time”, either explicitly or implicitly, then time would not be necessary. To investigate this, this essay looks at some current models of how the brain computes functions like these.
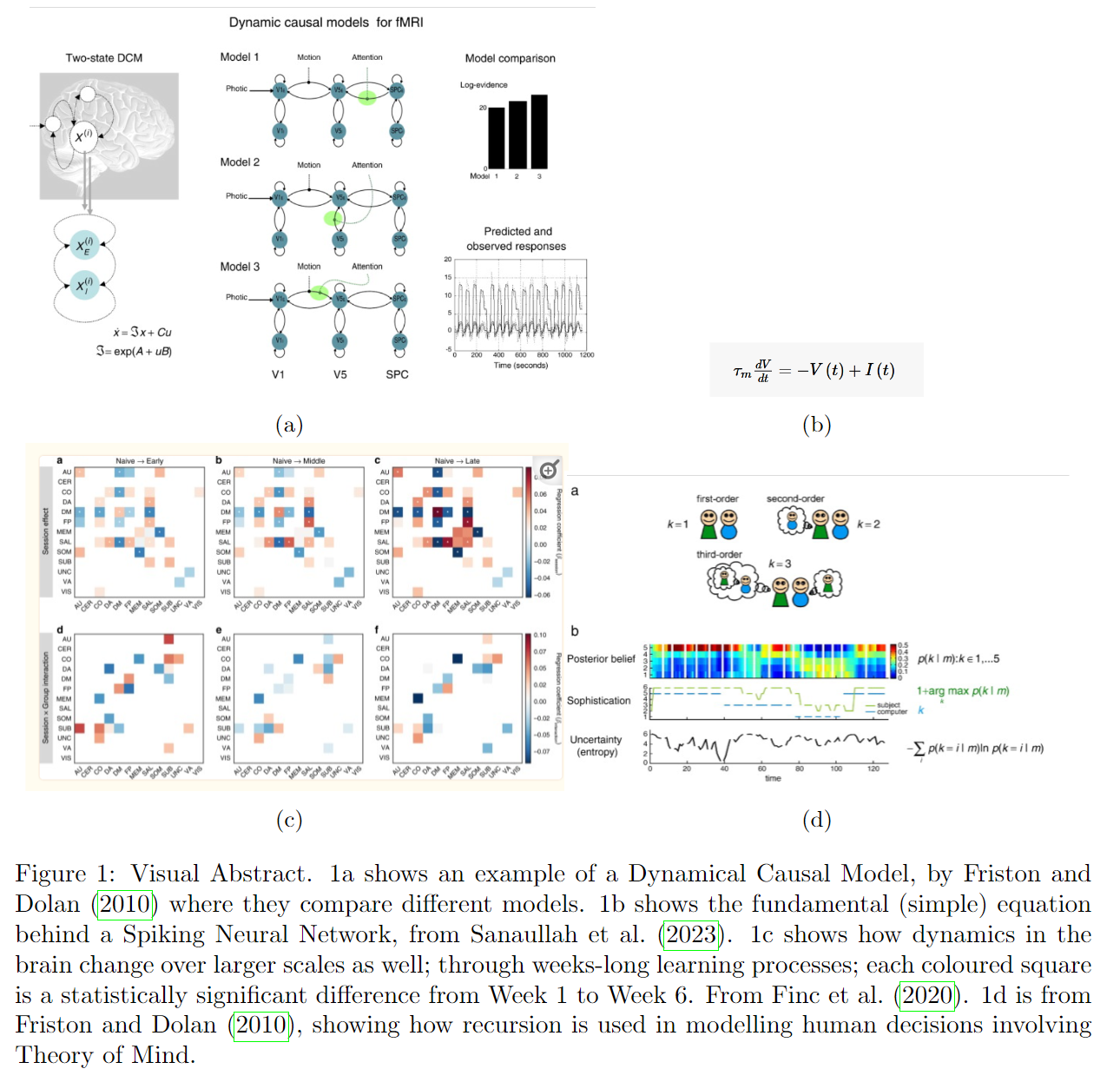
The first is similar to what Friston and Dolan (2010) calls neuronal dynamics. They survey the literature and find dynamic causal modelling to be appropriate, which is a framework within one uses differential equations to create models which, in effect, have “continuous time”. From this, we can infer that computational neuroscientists prefer models that include this time-aspect of neuronal dynamics.
If we turn to other avenues in computational neuroscience, we find that spiking neural networks have been gaining attention, “due to their ability to simulate the behaviour of biological neurons in the brain” (Sanaullah et al., 2023). Spiking neural nets are a type of neural net that incorporates discrete time steps into its computations. Once again, we see time pop us as, if not needed, then at the very least a useful modelling tool.
As for the second, Finc et al. (2020) analyses fMRIs of people who are new at a task, and how their brain adaptively changes over the course of 6 weeks spent gaining experience and expertise in that task. They find that, after training, the pattern of integration and recruitment of large-scale systems in the brain had changed.
In their review, Friston and Dolan (2010) concludes that, with certain relaxing assumptions, many, if not most, computational formulations of brain function “can usually be reduced to some form of prediction error”. This view of prediction error is close-to-inherently time-based, as the prediction error is built through learning from previous experience - previous time-steps.
An additional component, echoing through all of the above, and on full display in the work of cognitive scientist Douglas Hofstadter (e.g. Hofstadter (2007), is the idea of recursion and reflection; the mind is the way it is because it is continually reflecting its previous output. This too requires the idea of time in some way; for there to be a “previous” output there must in some sense be time.
The evidence showed here is not evidence that incorporating time-based dynamics might be necessary or sufficient. But it does show some ways in which modelling time is \textit{extremely useful}. From neuronal dynamics, learning, recursion, all of these depend on time in some way, and researchers in these fields use time-dependent dynamics, either through recurrent activity, online learning, or offline growing more resources devoted to specific problems, to model these problem areas. This is some evidence that time might be needed.
References
Finc, K., Bonna, K., He, X., Lydon-Staley, D. M., K¨uhn, S., Duch, W., & Bassett, D. S. (2020). Dynamic reconfiguration of functional brain networks during working memory training. Nature Communications, 11 (1), 2435. https://doi.org/10.1038/s41467-020-15631-z Friston, K. J., & Dolan, R. J. (2010). Computational and dynamic models in neuroimaging. NeuroImage, 52 (3), 752–765. https://doi.org/10.1016/j.neuroimage.2009.12.068 Hofstadter, D. R. (2007). I am a strange loop. Basic Books. Sanaullah, Koravuna, S., R¨uckert, U., & Jungeblut, T. (2023). Exploring spiking neural networks: A comprehensive analysis of mathematical models and applications. Frontiers in Computational Neuroscience, 17. https://doi.org/10.3389/fncom.2023.1215824
Essay 3
Essay Topic: Perceptual illusions might be a necessary feature in brains and artificial neural networks.
This essay will argue that perceptual illusions are not a necessary feature of visual perception systems, either in brains or artificial neural networks (ANNs), but that they might be \textit{likely} under certain constraints. I will draw on evidence, premises, and arguments from evolutionary theory and computational complexity theory. This essay is woefully incomplete and asks the reader to take several things on faith, which would ideally be presented either with argument or evidence, had I more time and space. I have annotated the relevant passages in the text with a “*”.
First, because ANNs are approximators of continuous functions (Funahashi, 1989), we must first ask; is it required that all continuous functions that compute visual perception be subject to these illusions? That seems like an arbitrary assumption to make, so for this essay I will assume$^*$ the opposite; that there exists a function for visual perception that avoids such visual illusions.
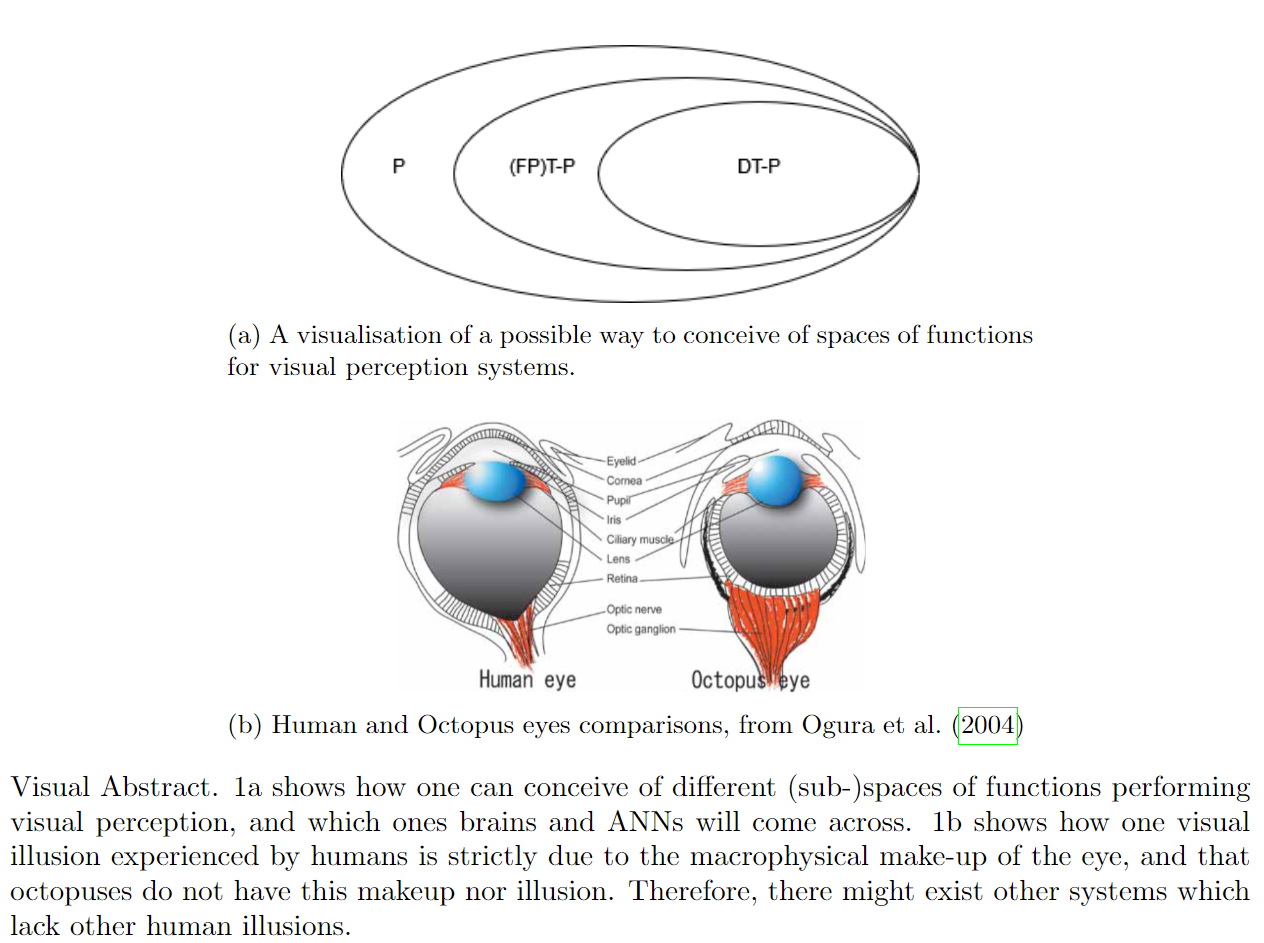
But if this is the case, why do our brains and ANNs exhibit these illusions? Consider the space of possible functions that compute visual perception. Let us call this space P. In reality, due to resource constraints, any cognitive system created could be computing only a subset of the functions in P. Let us call this subset “(FP)T-P”, for (Fixed Parameter) Tractable-Perception functions (Van Rooij, 2008). In addition to this, the visual perceptive systems in brains (and ANNs$^*$) have another constraint; they are reached in an incremental process. For brains; evolution, and ANNs; engineering (of the nets themselves) plus the backpropagation training loop. Let us call this subset DT-P, for (Fixed Parameter) Developmentally Tractable-Perception functions. See figure 1a for an illustration.
What might we expect functions in this subset to look like? From evolutionary theory we know that complex functions are built on simpler ones$^$. For visual systems, biologists have found that photoreceptive capabilities are adaptive in a wide range of environments Gehring, 2014. From there, a long series of small local improvements have led to the eyes of current species, which were reached from convergent evolution$^$, and are close to locally optimal$^*$ for their evolutionary environment.
However, some specifics of the visual system are just spandrels or “mistakes” in the evolutionary history. One standout example is the blindspot. Human retinas contain a spot where the optic nerve passes through the retina to the next part of the visual system, while the visual sensory organ of cephalopods like octopuses do not. I refer to Baden and Nilsson (2022) and Ogura et al. (2004) for a full explanation, and figure 1b for a visualisation. The point to make here is that systems which create certain perceptual illusions - like those begat by the blindspot - exist in DT-P alongside other systems which do not. And we cannot be sure which other systems might exist in DT-P.
So while we can engineer systems that do not have these perceptual illusions, brains - and ANNs - will stumble into local optima at several different optimization levels which lead them to exhibit these illusions, due to the evolutionary nature of their development. It is likely possible, but not guaranteed, to “engineer out” these perceptual illusions, but it will be difficult - akin to “remaking” the human visual system in the image of a cephalopod’s.
References
Baden, T., & Nilsson, D.-E. (2022). Is our retina really upside down? Current Biology, 32 (7), R300–R303. https://doi.org/10.1016/j.cub.2022.02.065 Funahashi, K.-I. (1989). On the approximate realization of continuous mappings by neural networks. Neural Networks, 2 (3), 183–192. https://doi.org/10.1016/0893-6080(89)90003-8 Gehring, W. J. (2014). The evolution of vision. WIREs Developmental Biology, 3 (1), 1–40. https://doi.org/10.1002/wdev.96 Ogura, A., Ikeo, K., & Gojobori, T. (2004). Comparative Analysis of Gene Expression for Convergent Evolution of Camera Eye Between Octopus and Human. Genome Research, 14 (8), 1555–1561. https://doi.org/10.1101/gr.2268104 Van Rooij, I. (2008). The Tractable Cognition Thesis. Cognitive Science, 32 (6), 939–984. https://doi.org/10.1080/03640210801897856